Addressing fundamental questions about the assembly and diversity of ecological communities
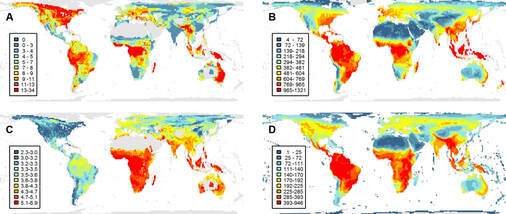
Studying how diverse ecological communities are assembled is challenging because most time series of community change are short (years to decades) and cover very limited spatial extents (metres to a few kilometres). This has inhibited our understanding of how biodiversity is generated and regulated as this requires also considering processes operating over macroevolutionary timeframes, such as speciation, dispersal, niche evolution and extinction. Our research is developing and applying phylogenetic approaches to model these dynamic process and to test how communities are formed and diversity is maintained. Much of our work has focussed on birds, which owing to the availability of geographic, phylogenetic and phenotypic data provide a model system. Our results have revealed how competition between species, dispersal and the historical timing of speciation events combine to limit the combination and number of sympatric species (Pigot et al 2018, Reijenga et al 2021), and how higher energy availability in the tropics facilitates the denser packing of ecological niche space (Pigot et al 2016). Our research has shown how this packing of ecological niche is a predictable process, with lineages on different continents (and that are thus freed from competition), repeatedly converging in their morphology to fill the same ecological niches (Pigot et al 2020). With a better understanding of how species are packed within communities and how their ecological niches and thus functions (e.g. seed dispersal, predation etc) have evolved, we are now developing improved models of how the functioning of ecosystems is responding to global change.
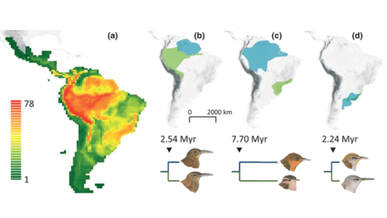
Top: examples of sister species of ovenbird of varying age, showing the different geographical stages in the build up of sympatric diversity, from speciation in allopatry to range expansions leading to parapatry and eventually sympatry.
Middle: hotspots of species richness (bottom right map) coincide with hotspots of sympatry between sister species (top left map) which are strongly predicted by energy availability (top right map) and species age (bottom left map).
Studying how diverse ecological communities are assembled is challenging because most time series of community change are short (years to decades) and cover very limited spatial extents (metres to a few kilometres). This has inhibited our understanding of how biodiversity is generated and regulated as this requires also considering processes operating over macroevolutionary timeframes, such as speciation, dispersal, niche evolution and extinction. Our research is developing and applying phylogenetic approaches to model these dynamic process and to test how communities are formed and diversity is maintained. Much of our work has focussed on birds, which owing to the availability of geographic, phylogenetic and phenotypic data provide a model system. Our results have revealed how competition between species, dispersal and the historical timing of speciation events combine to limit the combination and number of sympatric species (Pigot et al 2018, Reijenga et al 2021), and how higher energy availability in the tropics facilitates the denser packing of ecological niche space (Pigot et al 2016). Our research has shown how this packing of ecological niche is a predictable process, with lineages on different continents (and that are thus freed from competition), repeatedly converging in their morphology to fill the same ecological niches (Pigot et al 2020). With a better understanding of how species are packed within communities and how their ecological niches and thus functions (e.g. seed dispersal, predation etc) have evolved, we are now developing improved models of how the functioning of ecosystems is responding to global change.
Top: examples of sister species of ovenbird of varying age, showing the different geographical stages in the build up of sympatric diversity, from speciation in allopatry to range expansions leading to parapatry and eventually sympatry.
Middle: hotspots of species richness (bottom right map) coincide with hotspots of sympatry between sister species (top left map) which are strongly predicted by energy availability (top right map) and species age (bottom left map).
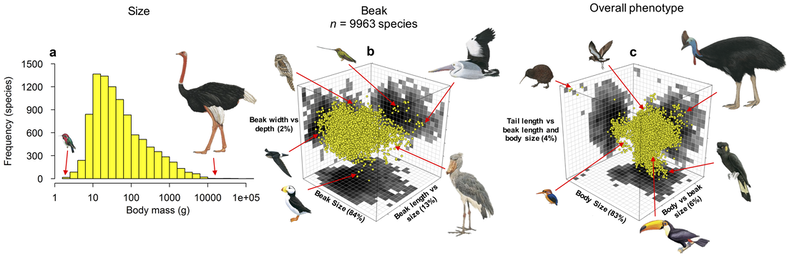
Understanding and predicting risks to biodiversity in the Anthropocene
Ecological communities are changing at unprecedented rates due to human activity. The transport of species around the world is resulting in an unprecedented mixing of long isolated biotas, the expansion of human land uses is providing novel selective challenges and opportunities for species, with climate change driving many environments into states with closest analogues deep in Earths history. Our research seeks to understand how these anthropogenic drivers are altering Earths ecosystems (e.g. Lovell et al 2021).
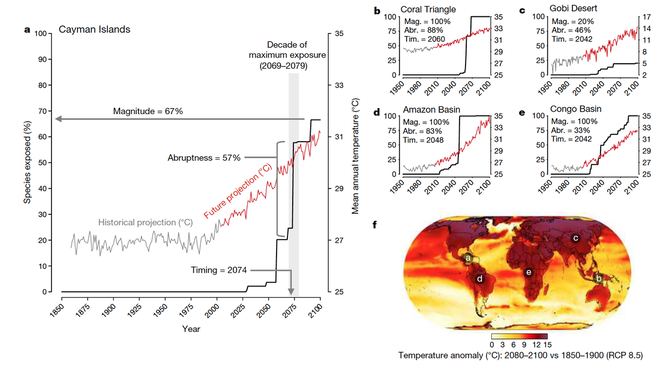
Just as it is difficult to infer how communities have dynamically assembled from only looking at a single snapshot of community structure at the present day, our understanding of how biodiversity will change into the future has also been limited by focussing on one or a few snapshots (e.g. the year 2070 or 2100). We have been developing a new framework for modelling how risks to biodiversity are projected to change over much finer temporal resolutions (i.a. monthly-annually), the biodiversity 'horizon profile' (Trisos et al 2020). Horizon profiles describe the cumulative proportion of species in an assemblage (or populations across a species' geographic range) that are exposed to conditions beyond their niche limits over time. This provides more information on risks to biodiversity that simply the magnitude of exposure (i.e. proportion exposed at any single snapshot), including when and how abruptly exposure will occur.
We have been applying this framework to terrestrial vertebrates and marine organisms globally, finding that exposure of species within assemblages to dangerous climate conditions (i.e. beyond the range of temperatures they have previously experience) is likely to occur abruptly, with multiple species exposed near simultaneously across wide geographic areas, suggesting that species and ecological communities may be at risk of abrupt collapse under global warming. Maps of projected biodiversity exposure can be viewed here (link).
Our models have been used as one of the key biodiversity risk metrics in the IPCC 6th Assessment Report and we are now working with conservation organisations, including South African National Parks, to use our projections to inform protected area management and climate adaptation.
Ecological communities are changing at unprecedented rates due to human activity. The transport of species around the world is resulting in an unprecedented mixing of long isolated biotas, the expansion of human land uses is providing novel selective challenges and opportunities for species, with climate change driving many environments into states with closest analogues deep in Earths history. Our research seeks to understand how these anthropogenic drivers are altering Earths ecosystems (e.g. Lovell et al 2021).
Just as it is difficult to infer how communities have dynamically assembled from only looking at a single snapshot of community structure at the present day, our understanding of how biodiversity will change into the future has also been limited by focussing on one or a few snapshots (e.g. the year 2070 or 2100). We have been developing a new framework for modelling how risks to biodiversity are projected to change over much finer temporal resolutions (i.a. monthly-annually), the biodiversity 'horizon profile' (Trisos et al 2020). Horizon profiles describe the cumulative proportion of species in an assemblage (or populations across a species' geographic range) that are exposed to conditions beyond their niche limits over time. This provides more information on risks to biodiversity that simply the magnitude of exposure (i.e. proportion exposed at any single snapshot), including when and how abruptly exposure will occur.
We have been applying this framework to terrestrial vertebrates and marine organisms globally, finding that exposure of species within assemblages to dangerous climate conditions (i.e. beyond the range of temperatures they have previously experience) is likely to occur abruptly, with multiple species exposed near simultaneously across wide geographic areas, suggesting that species and ecological communities may be at risk of abrupt collapse under global warming. Maps of projected biodiversity exposure can be viewed here (link).
Our models have been used as one of the key biodiversity risk metrics in the IPCC 6th Assessment Report and we are now working with conservation organisations, including South African National Parks, to use our projections to inform protected area management and climate adaptation.
Identifying the limits to the resilience of biodiversity
The thermal tipping points that our models reveal suggest that abrupt collapses of species and communities should be expected even in the absence of the kind of positive ecological feedbacks typically invoked to explain why populations and ecosystems can rapidly disintegrate. A key focus of our research (the 'tipping points' project) is now to understand how biodiversity, both within and across species, may buffer communities from abrupt collapse. Using butterflies as a model system, we are combining experiments of species thermal physiology with measurements and models of body temperature and microclimates, to test how organism behaviour to exploit fine scale climate heterogeneity in space and time, either tempers, delays or amplifies risks of abrupt collapse as species approach and eventually exceed their thermal limits.
The thermal tipping points that our models reveal suggest that abrupt collapses of species and communities should be expected even in the absence of the kind of positive ecological feedbacks typically invoked to explain why populations and ecosystems can rapidly disintegrate. A key focus of our research (the 'tipping points' project) is now to understand how biodiversity, both within and across species, may buffer communities from abrupt collapse. Using butterflies as a model system, we are combining experiments of species thermal physiology with measurements and models of body temperature and microclimates, to test how organism behaviour to exploit fine scale climate heterogeneity in space and time, either tempers, delays or amplifies risks of abrupt collapse as species approach and eventually exceed their thermal limits.


Research sites in the Sierra de Guadarrama where we are conducting experiments and field work in collaboration with Rob Wilson, Jon Bridle and Ilya Maclean to understand tipping points in butterfly communities across thermal gradients in space and time
